

Přechod na souš
Devon byl tradičně považován za "období ryb". Během devonu se uskutečnily dramatické proměny všech hlavních linií ryb. V devonu se také poprvé objevují lalokoploutvé ryby stejně jako paprskoploutvé ryby (dnes nejrozsíšřenější skupina ryb vůbec). Lalokoploutvé ryby dosahují vrcholu své diverzity během devonu a daří se jim také během následujícího období - karbonu. Devon je skutečně velmi zajímavé období v historii naší Země a hlavně v historii nás samých. Jedná se totiž o období, kdy se objevili první suchozemštví obratlovci.
Příležitost volá
Když se dnes podíváme na obrovskou diverzitu plazů, savců a ptáků, tak musíme žasnout nad tím, jak velkou cestu museli naši předci překlenout, když se dostávali z vody na souš. Změn bylo zapotřebí opravdu hodně. Ať už se jedná o ty velmi nápadné, jako je změna stavby těla nebo kostry končetin. Naši předci se museli naučit dýchat vzdušný kyslík a zabezpečit své tělo proti velmi nepřátelskému prostředí, které na ně na souši čekalo. To jak k takové proměně mohlo dojít provází i otázka proč k tomu došlo.
 |
| Obr. 1 Dunkleosteus |
Důvodů proč přejít na souš bylo hned několik. Abysme si však tyto důvody mohli lépe představit je nutné podívat se na ekologické poměry, které zde panovaly právě během devonu. Příležitosti se jen nabízely. Během pozdního devonu se objevila celá řada nových, do té doby nepoznaných, biotopů. Souš byla zaplavena rostlinami a objevují se i první stromy. Rozsáhlá pozemská vegetace stabilizovala břehy a spodní sedimenty. To přímělo různé živočichy přejít z hlubokých vod do mělčin, které periodicky zaplavovaly přilehlé mokřady. Hluboké kanály, močály, zátoky nebo periodicky se zaplavující planiny. To vše je v devonu nové a to vše nabízí celou řadu příležitostí pro rozvoj života. Adaptace na tyto mělké vody a bažiny jednou dovolí tetrapodům expandovat na suchou zemi. 1
Zatímco na souši rostou jen rostliny a poletuje tu hmyz, ve vodě se to hemží obrovskými dravci jako byl například devítimetrový Dunkleosteus nebo sedmimetrový Rhizodus. 1 Zarostlé mělčiny a mokřady znamenaly nové zdroje potravy a zřejmě také zabezpečovaly útočiště pro mladé stejně jako byly bezpečné pro dospělé jedince. Souš je v té době klidné mírumilovné místo pro ryby však zatím nedostupné. Nicméně různé zátoky a řeky se pro mnohé ryby stávají bezpečným útočištěm, kde se mezi kořeny a padlými stromy mohou bezpečně pohybovat.
 |
| Obr. 2 Periodicky se zaplavující tůně |
Nově vzniklé periodicky se zaplavující pláně mohou uvěznit celé hejno ryb na poměrně dlouhou dobu. V takto stojatých vodách kyslík rychle ubývá a jasnou výhodou je možnost dýchat kyslík vzdušný. Něco podobného dnes dokáží dvojdyšné ryby. Proto schopnost vydržet déle na vzduchu byla výhodou pro ryby, které chtěly uniknout predátorům ukrytím se v bažinách a močálech stejně jako pro ryby nedobrovolně uvězněné ve stojatém jezírku, které zbylo na místě předchozí povodně. 1
Morfologické změny
Molekulární a morfologické analýzy ukazují, že k oddělení tetrapodů od ryb došlo někdy ve spodním karbonu nebo ve svrchním devonu. Pro přeměnu ploutve v končetinu s prsty bylo zapotřebí mnoho dalších důležitých změn. Především muselo dojít ke zvětšní hrudního a pánevního kostního pletence, vzniku kosti křížové, spojení žeber s obratli, vznik krku, změna lebečních proporcí, změna smyslových systémů, redukce žaberního oblouku a redukci řitní a hřbetní ploutve.
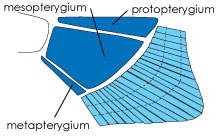 |
| Obr. 3 Tři bazální elementy žraločí ploutve |
Vývoj končetin z nejprimitivnějších ploutví můžeme sledovat také na základních elementech kostry končetin. Významnou roli při vzniku končetin hrají tři bazální elementy (bazalia), ze kterých u ryb vybíhají paprsky ploutví (radialia). U nejprimitivnějších čelistnatých obratlovců (Gnathostomata - paryby) jsou přítomny tři bazální elementy: protopterygium, mesopterygium a metapterygium. 6
V dalším vývoji dochází k rozdělení ryb do dvou skupin. První skupinu tvoří Actinopterygii, kam řadíme také většinu dnes známých ryb - ryby kostnaté (Teleostei). U této první skupiny dochází prakticky ke ztrátě metapterygia. Další vývojovou větev tvoří skupina Sacropterygii. Sem patří lalokoploutvé ryby, které se objevují poprvé v devonu. Kromě těchto ryb se však do této skupiny zahrnují také všichni tetrapodi. U těchto ryb došlo k procesu zcela opačnému. Došlo ke ztrátě protopterygia a mesopterygia, takže paprsky ploutví vybíhají jen z kosti metapterygia. A právě na tuto bazální kost navazují u pozdějších druhů prsty. 6
Metapterygium ovšem mohlo hrát významnou roli ve vývoji končetin i z jiného důvodu. Někde na přechodu mezi parybami a rybami dochází k pootočení metapterygia o 90°. Ve skutečnosti se pochopitelně tato kost nepootočí, ale v průběhu embryogeneze, právě když vzniká a roste končetinový pupen, dělí se buňky v zadní (posteriorní) části budoucího metapterygia více, než v přední (anteriorní), takže zadní část se jakoby vytočí ven, v distálním směru vznikající končetiny. Zajímavé je, že u těch druhů, kde je hlavní osa metapterygia ještě shodná s osou těla, nebyla detegována v končetinovém pupenu exprese genu Sonic hedgehog, která se vždy objevuje tam, kde je již metapterygium natočené směrem ven z těla. To by mohlo znamenat, že Sonic hedgehog je tím genem, který odstartoval vývoj, vedoucí přes větší autonomii celé končetiny k takovému jejímu typu, ve kterém hraje zásadní roli právě metapterygium, tedy ke kráčivé končetině suchozemských obratlovců. 6
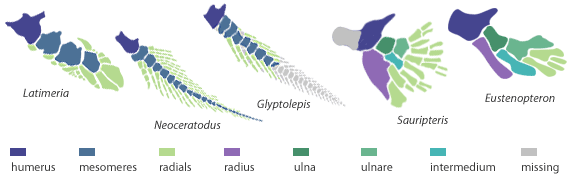 |
| Obr. 4 Anatomie ploutví žijících a vymřelých lalokoploutvých ryb |
Už jsme si řekli, že se v devonu objevují poprvé lalokoploutvé ryby - skupina Sacropterygia. Ve skutečnosti dochází k rozvoji hned čtyř skupin lalokoploutvých ryb. Jedná se o skupinu dvojdyšných ryb Dipnoi (Neoceratodus viz Obr.), dále o vymřelou skupinu Porolepiformes (Glyptolepis viz Obr.), jejichž ploutve sdílí celou řadu anatomických podobností se skupinou Dipnoi. Jedná se hlavně o výraznou centrální osu ploutve. Další skupinou, která má dnes už jen dva zástupce je skupina Actinistia (Latimeria viz Obr.). Dnes tuto skupinu reprezentují jen dva druhy latimérií. Poslední skupinou byla skupina Rhipidistia (neboli Choanata), která sice vymřela, ale nechala nám tu dědictví v podobě tetrapodů. Ano... Právě z této poslední skupiny se podle všeho dnešní suchozemští obratlovci vyvinuli. Tuto skupinu lze rozdělit na dvě podskupiny - Rhizodontida (Sauripteris viz Obr. 4) a Osteolepidida (Eusthenopteron viz Obr. 7, Tiktaalik a Panderichthys viz Obr. 8).
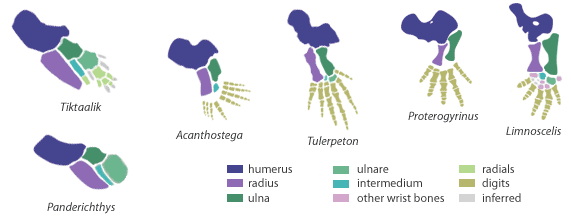 |
| Obr. 5 Dvě ploutve Rhipidistií a čtyři končetiny tetrapodů |
Z obrázků je patrné, že protažené metapterygium se vlastně stává hlavní osou končetiny. Všechny kostěné elementy, které z něj vybíhají směrem k přednímu konci těla (anteriorně) jsou pak označovány jako preaxiální, ty které jdou směrem dozadu (posteriorně) jako postaxiální. Prsty u tetrapodů jsou tak evoluční novinkou v podobě postaxiálních elementů.
 |
 |
| Obr. 6 Fosílie stromu Archaeopteris a jeho rekonstrukce | |
Porovnámeli stavbu ploutví se stavbou končetiny suchozemských obratlovců, tak je jasné, že nejblíže k ní mají právě lalokoploutvé ryby ze skupiny Rhipidistia. Není snad náhodou, že tyto ryby obsahují celou řadu dalších podobností s tetrapody. Obrázek ukazuje porovnání ploutví dalších ryb ze skupiny Rhipidistia a z řádu Osteolepiformes s končetinami prvních tetrapodů. Jediný rozdíl mezi končetinou a ploutví je v kompletní ztrátě lepidotrichií (paprskovitých kostí vycházejících z hlavních kostí ploutve) a jejich nahrazením prsty.
Skupina Rhipidistia postupně přichází o hřbetní a břišní ploutve a zůstanou jen dvě párové ploutve v mnohém podobné končetinám. Ovšem stále bez prstů. U celé řady devonských ryb sice vidíme jakousi obdobu prstů (např. Sauripteris, Tiktaalik,...), ale k čemu vůbec sloužily tyto v podstatě mohutné kostěné ploutve u ryb? V devonu se rozšiřují lesní porosty a jedním z dominantních rodů je rod Archaeopteris. Jedná se o velmi rozšířený rod. Tento druh strumu má zajímavou vlastnost. Když opadávají jeho zvláštní listy, tak vlastně upadnou s celou větví. V této spleti větví, které dopadnou na dno nějaké zátoky nebo močálu je výhodou mít mohutné a pohyblivé ploutve. Jejich funkci si můžeme představit na dnes žijícím druhu, který však s těmito rybami nemá nic společného. Jedná se o mořské ďasy, kteří své ploutve zdatně používají jakoby to byly končetiny s prsty. Pohybují se pomocí nich na kamenitém dnu nebo na korálových útesech.
Život v devonských vodách
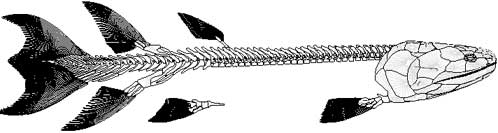 |
| Obr. 7 Eusthenopteron |
Typickým zástupcem skupiny Rhipidistia byl Eusthenopteron. Tato ryba je známa ze stovek dochovaných exemplářů, které byly nalezeny v oblasti Miguasha kanadského Quebecu v hornině staré asi 364-370 milionů let. Paleontolog Erik Jarvik vytvořil 3D model tohoto druhu podle výborně zachovalého exempláře. Eusthenopteron byl asi 1,5 metrů dlouhý, měl ještě zachovalou jak řitní tak i hřbetní ploutev a jeho ploutve v mnohém připomínaly nohy bez prstů. Měly podobnou stavbu jako u všech suchozemských obratlovců. Měl postaxiální kostnaté výběžky připomínají vzdáleně prsty. Tato ryba se však podobala skutečně ještě spíše rybám než obojživelníkům. I když měla některé znaky, které se objevují právě až u tetrapodů.
 |
| Obr. 8 Panderichthys rekonstrukce |
Dalším zástupcem této zajímavé skupiny ryb byl rod Panderichthys. Ten byl nalezen v Litvě v hornině staré 378 milionů let a podle místa nálezu je pravděpodobné, že žil ve sladkých vodách.. Jsou známy dva druhy. Panderichthys stolbovi, který je znám pouze podle fragmentů lebky a Panderichthys rhombolepis jehož fosilní materiál je mnohem bohatší. Původně byl Panderichthys klasifikován jako primitivní tetrapod, ale po nálezu jeho končetin muselo být jeho zařazení překlasifikováno.
Panderichthys byl asi 90-130 cm dlouhý, měl sploštělou hlavu velmi podobnou tetrapodům, která se od úzkého čenichu rozšiřovala směrem dozadu. Mezilebeční kloub, který je charakteristický pro lalokoploutvé ryby, byl odstraněn z povrchu lebky, ale je stále umístěn uvnitř lebky. Povrchová struktura kostí na horní části lebce a na lících je podobná více primitivním tetrapodům než lalokoploutvým rybám.
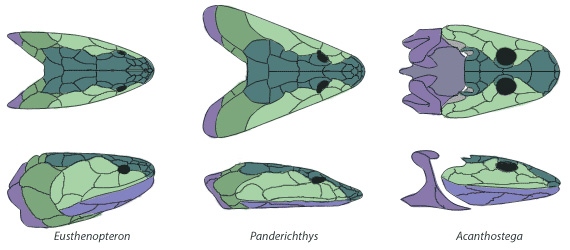 |
| Obr. 9 Porovnání lebek u lalokoploutvých ryb a prnvího obojživelníka |
I z ostatních částí kostry lze vypozorovat kvalitu tohoto přechodného článku. Chybí jak hřbetní tak i řitní ploutev a ocas se více podobá ocasu prvních tetrapodů než tomu u lalokoploutvých ryb. Rameno ukazuje několik tatrapodům podobných vlastností. Je delší než u všech ostatních lalokoploutvých ryb. Na druhou stranu spodní část končetiny se končetinám tetrapodů moc nepodobá. Je to ploutev, která obsahuje množství dlouhých tenkých paprsčitých kostí. Kosti zadních končetin byly po pečlivé preparaci získány z fosilního materiálu poměrně nedávno 2. Ukazuje se, že jsou složeny z mohutných kostí, nemají prsty, ale pouze tenké kostěné paprsky. Obratle jsou podobné více obratlům primitivních tetrapodů.
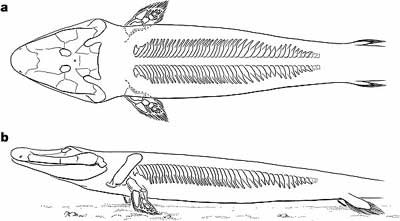 |
| Obr. 10 Tiktaalik |
Před nedávnem byl objeven další předchodný článek od ryb na cestě k obojživelníkům. Tento tvor dostal název Tiktaalik roseae a jeho existence byla předpovězena paleontology před několika lety. Byl objeven v oblasti Nunavut v arktické Kanadě.
Tiktaalik byl poměrně velkou rybou. Nalezený exemplář mohl podle odhadů dosahovat délky až jeden metr a podle kostry lze usuzovat, že někteří jedinci mohli dorůstat až dvoumetrové délky. V mnohém připomíná dnešní krokodýly s širokou a spolštělou lebkou, s nozdrami vyčnívajícími z lebky, s očima posazenýma nahoře na lebce a s kostěnými šupinami. Kostra jednoznačně ukazuje na to, že tento tvor trávil hodně času v mělkých vodách.
Tiktaalik ukazuje celou řadu přechodných prvků. Jeho lebka mnohem více připomíná lebku prvních obojživelníků než lebka Panderichthyse. Dokonce i jednotlivé kosti na lebce souhlasí více s kostmi obojživelníků než s kostmi předchozích ryb. Na druhou stranu spodní čelist je více podobná té u Panderichthyse. Stejně jako všechny ryby má i Tiktaalik dobře vyvinuty vnitřní žábry. Důležitým znakem je také ztráta kostěného "límce" spojujícím lebku s ramenním pletencem. Jinými slovy to znamená, že Tiktaalik měl krk jako všichni obojživelníci. Jeho žebra jsou dokonce více vyvinuta než žebra u Acanthostegy a jeho páteř je také v mnohém unikátní.
Do této chvíle jsme mluvili v podstatě stále o rybách. Nejstarším objeveným obojživelníkem je však až Acanthostega gunneri. První fosíle tohoto druhu byla objevena už v roce 1933 ve východním Grónsku. Jednalo se však pouze o horní část lebky. Erik Jarvik ji popsal a pojmenoval až v roce 1952. Další nálezy tohoto druhu byly objeveny během expedice v roce 1970. Zlom nastal až v roce 1987, kdy během expedice vedené Clackem a Perem Ahlbergovými byly objeveny výjimečně dobře zachovalé zkameněliny několika jedinců.
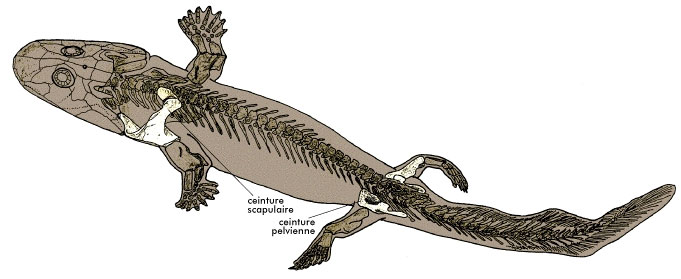 |
| Obr. 11 Acanthostega |
Do té doby si paleontologové mysleli, že se u tetrapodů vytvořily končetiny pro pohyb po souši. Acathostega však přinesl odpověď poněkud jinou. Je zajímavé, že ačkoliv měl Acanthostega dobře zformované prsty, neměl žádné kotníky ani zápěstí. Měl poměrně dlouhé kosti na končetinách, které však nebyly schopny unést větší váhu na souši. Stejně tak byla formována i kyčel. Ta totiž byla velmi slabě připojena k páteři a nebyla tak schopna unést celou váhu při pohybu po souši. Dokonce ani pevnější spojení kyčle k páteři by moc nepomohlo. Jeho páteř byla totiž podobná spíše hřbetní struně než do sebe zapadajícím obratlům. Byla stavěna pro odolávání mechanickým tlakům při plavání, ale pro pohyb po souši byla nevyhovující. Navíc žebra tohoto živočicha byla příliš krátká na to, aby chránila vnitřní orgány při pohybu po souši. Kromě toho měla Acanthostega dlouhý ocas ne nepodobný ocasním ploutvým lalokoploutých ryb. Tento ocas jednoznačně sloužil při pohybu ve vodě.
 |
| Obr. 12 Rekonstrukce Acanthostegy |
Primární nebo dokonce výhradní vodní způsob života u Acantostegy dokládají i vnitřní žábry podobné rybím. Všichni obojživelníci od karbonu až dodnes mají vnější žábry. Acanthostega měl však žábry vnitřní stejně jako je tomu u ryb. To dokládá kostěný žaberní oblouk a postbrachiální destička na přední hraně ramenního pletence. Další důkaz vodního způsobu života je kost, která se u tetrapodů později vyvine v jednu z uších kůstek (třímenk). U Acanthostegy je podobná rybímu ekvivalentu této kosti. Jeho malé rybám podobné nozdry byly uzpůsobeny pravděpodobně pouze k čichání pod vodou. Vzduch Acanthostega nejspíš polykala.
Ačkoliv má Acanthostega mnohé rybí znaky její končetiny se podobající více nohám než ploutvím. Tyto končetiny ovlivnily naše nazírání na evoluci tetrapodů. Byla spíše víra v to, že první tetrapodi měli pět prstů. Acanthostega má však na přední končetině osm a na zadní nejméně osm prstů. Jak se ukázalo ještě nejméně dva další známé druhy prvohorních obojživelníků měli více než pět prstů (Ichthyostega a Tulerpeton).
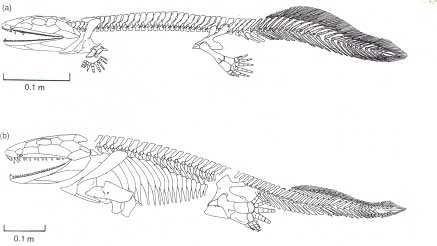 |
| Obr. 13 Porovnání koster Acanthostegy a Ichthyostegy |
Acanthostega tedy se svými rybími charakteristikami nebyla schopna pohybu po souši. Žila zřejmě v mělkých vodách (mokřady, potoky, záplavové planiny), které obsahovaly vodní vegetaci a její končetiny jí sloužily při pohybu po dně nebo je využívala jako pádla při plavání. Acanthostega byla dlouhá asi 60 cm a žila před asi 360 miliony let.
Dalším raným obojživelníkem byl Ichthyostega stensioei. Ichthyostega byla nalezena ve východním Grónsku stejně jako Acanthostega. Byl to první vědě známý devonský tetrapod. Geologická vrstva (stáří asi 360 milionů let), ve které byly fosíle nalezeny napovídá tomu, že tento živočich žil pravděpodobně v potoce. Původně byl fosilní materiál pokládán za tři druhy rodu Ichthyostega a jeden druh Ichthyostegopsis. Dnes jsou však všechny čtyři fosílie přiřazeny pouze k jednomu druhu Ichthyostega stensioei. Podle posledních studií se jednalo o více vodního živočicha než suchozemského. Každopádně byl pro suchozemskou chůzi přizpůsoben lépe než Acanthostega.
 |
| Obr. 15 Fosílie Ichthyostegy |
Ichthyostega byl poměrně velký asi jeden a půl metru dlouhý obojživelník s nápadně tlustým tělem. Původně byl pokládán za přechodnou formu mezi rybami a karbonskými tetrapody, nicméně jeho rybám podobná lebka jé více suchozemského typu než rybího ve srovnání s Acanthostegou.
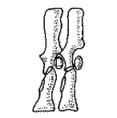 |
| Obr. 14 Obratle Ichthyostegy |
Stejně jako u Acanthostegy je páteř spíše založena na principu hřbetní struny než na principu do sebe zapadajících avšak flexibilních obratlů, jako je tomu u pozdějších tetrapodů. Páteř však už vypadá na to, že dokáže podpírat tělo při pohybu po souši. Ocas Ichthyostegy je velmi podobný rybímu avšak kostěné výběžky jsou menší než u Acanthostegy. Nápadná je však stavba jeho hrudníku. Žebra jsou zploštělá, mohutná a překrývají z části jedno nebo více sousedících žeber. To vytváří pevnou schránku chránící tělo při pozemní chůzi. Zároveň z toho však vyplývá, že jeho pohyb při chůzi nebo plavání nemohl být tak flexibilní právě kvůli tomuto pevnému krunýři.
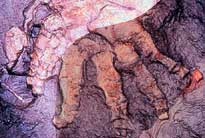 |
| Obr. 16 Zadní končetina Ichthyostegy |
Ramenní pletenec byl velmi podobný ramennímu pletenci Acanthostegy. Žádná známka po postbrachiální destičce nebyla nalezena, z čehož lze usuzovat, že Ichthyostega neměl vnitřní žábry. Přední končetiny jsou robustnější než u Acanthostegy, takže se zdá, že mohly podpírat část váhy těla. Ramenní pletenec byl poměrně flexibilní, ale loket dovoloval jen velmi malý pohyb směrem dopředu nebo dozadu. Kostra předního chodidla se nedochovala. Pánev byla velká a jak se zdá, tak i připojená k páteři. Zadní končetiny byly menší než přední a sloužily spíše k pádlování ve vodě než k chůzi. Koleno bylo poměrně flexibilní, avšak chybí kotník. Na zadních končetinách měl Ichthyostega sedm prstů.
Původní rekonstrukci, tak jak ji asi všichni známe, zobrazuje obrázek dole. Není však jasné, zda-li mohly být jeho zadní končetiny schopny takového postavení. Pravděpodobnější je, že je používal při plazení asi tak jak dnes lvouni. Jejich postavení tedy více odpovídá rekonstrukci úplně nahoře na prvním obrázku.
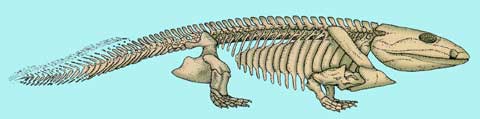 |
| Obr. 17 Původní rekonstrukce Ichthyostegy |
Posledním důležitým obojživelníkem, kterého zde popíšu je Tulerpeton curtum. Tulerpeton žil v pozdním devonu v Rusku v oblasti Tula. Nález je docela bohatý v porovnání s ostatními tetrapody dochovanými z té doby (avšak nedosahuje takové kvality jako u Ichthyostegy nebo Acanthostegy). Dochovaný materiál se skládá z téměř kompletního hrudního pletence, přední i zadní končetiny, z fragmentů lebky a pánve a množství břišních šupin.
Nejzajímavější na Tulerpetonu jsou však jeho končetiny. To z toho důvodu, že obsahují vyšší počet prstů než u všech ostatních tetrapodů (kromě Ichthyostegy a Acanthostegy). Z toho vyplývá, že pětiprstá končetina se vyvinula pravděpodobně až v karbonu. Prsty jsou relativně dlouhé a úzké z čehož se dá usuzovat na pozemní způsob života. Nicméně kotníky a způsob svalového uchycení stehenní kosti spíše vypovídá o tom, že tyto zadní končetiny byly lépe uzpůsobeny k pádlování ve vodě než k plazení po souši.
Ramenní pletenec je více robustní než u druhu Ichthyostega, což vypovídá o více pozemnímu než vodnímu způsobu života. Chybí zde i postbrachiální destička, takže Tulerpeton neměl vnitnřní žábry. Na druhou stranu jeho ramenní pletenec si ponechal horní část (anocleithrum), který má i Acanthostega, ale například Ichthyostega již tuto část nemá. Z mála fragmentů lebky lze říct, že Tulerpeton měl blíže ke karbonským obojživelníkům než výše zmíněným dvěma devonským druhům. Zatímco předchozí dva druhy byly objeveny ve vrstvách, které vypovídají o sladkovodním životě, Tulerpeton byl nalezen ve vrstvách, které ukazují na přímořskou oblast.
Tulerpeton není posledním obojživelníkem z tohoto období. Existuje celá řada mu podobných obojživelníků jako byli Densignathus, Hynerpeton, Jakubsonia, Metaxygnathus nebo Ventastega. Jedná se však o exempláře známé jen díky velmi fragmentovanému fosilnímu záznamu.
 |
| Obr. 18 Stopy raných tetrapodů z Austrálie z Nového Jižního Walesu |
Zajímavou otázkou jsou i vzácné nálezy z Austrálie. Většinu dokladů o evoluci tetrapodů známe prakticky jen ze severní polokoule. Ovšem existuje i několik dokladů o tom, že ryby opustily vodu i na polokouli jižní a to právě v Austrálii. Bohužel fosilní záznam nám o této události nic moc neřekne. Jedná se jen o jednu spodní čelist raného obojživelníka (Metaxygnathus) a několik stop. Tyto fragmety mohou ukazovat dvě věci. Buď, že došlo k vývoji obojživelníků dvakrát a nezávisle na sobě nebo celý vývoj probíhal na větším území než jsme se doposud domnívali. Austrálie od severní Evropy a Severní Ameriky nebyla v devonu zase tak nepřekonatelně daleko. Nicméně ji dělilo moře a cesta z jedné části světa na druhou by byla obtížná, nicméně ne nemožná.
Jak dýchat kyslík ze vzduchu
Tento poměrně bohatý materiál nám ukazuje cestu prvních ryb na souš a to z anatomického hlediska. Bohužel celá řada znaků, které dovolují obojživelníkům přežívat na suchu se ve fosilním záznamu nedochovává. Jedná se také o adaptaci, která dovoluje tetrapodům dýchat vzdušný kyslík. Jedná se o nejnápadnější adaptaci tetrapodů vůbec. Takové dýchání je samozřejmou nezbytností pro přizpůsobení se suchozemskému životu. Je to však také příznivé pro velká zvířata zkoušející využít mělké stojaté vody.
Naneštěstí je evoluce a původ plic nejasná. Plíce se pravděpodobně vyvinuly dvakrát nezávisle. Jednou u dvojdyšných ryb a podruhé u řádu Osteolepiformes potažmo u tetrapodů. Velmi ranné mořské dvojdyšné ryby pravděpodobně neměly funkční plíce, ale některé pozdně devonské a karbonské druhy si patrně vytvořily nějaký mechanismus pracující na principu polykání vzduchu, který jim dovoloval dýchat trochu vzduchu. Dýchání pomocí žaber je pak dále redukováno a dnes pokud některé žijící dvojdyšné ryby pokud nemají přístup k volnému kyslíku, tak se utopí.
První funkční plíce v linii tetrapodů nejsou známy. Je však známo, kdy zanikly interní žábry v linii Osteolepiformes. Ještě první tetrapodi jako byl Acanthostega měli vnitřní žábry. Jakýkoliv důkaz o vnitřních žábrech u ostatních pozdně devonských tetrapodů chybí nebo se nedochoval.
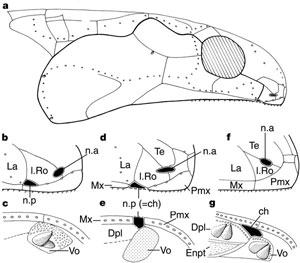 |
| Obr. 19 Vznik choany (b,c - Youngolepis; a,d,e - Kenichthys; f,g - Eusthenopteron) 4 |
Trápení nad linií důkazů spojených se vzdušným dýcháním zahrnuje i tzv. choanu. Je to struktura spojující vnější nozdry s dutinou ústní. Obsahuje také otvor, který vede do očních důlků přes malý kanálek. Choana je podle všeho homologická s primitivními párovými vnějšími nozdrami, které můžeme stále najít u mihulí, žraloků, latimérií a paprskoploutvých ryb. Tyto vnější nozdry slouží k čichání pod vodou, ale k dýchání už ne. Choana byla nalezena pouze u řádu Osteolepirofmes a pozdějších tetrapodů. Jejich přítomnost u rhizodontů však nebyla rozpoznána. Zajímavé je to, že tato struktura byla nalezena i u odvozených dvojdyšných ryb. Nicméně u těchto ryb chybí kanálek vedoucí do očních důlků.
Choana je základním kanálem, kterým moderní tetrapodi dýchají. Jejich funkce u prvních tetrapodů a lalokoploutvých ryb z řádu Osteolepiformes je příliš nejasná. Mohla být používána k dýchání vzdušného kyslíku dokonce i u plně vodních živočichů jako byl Eusthenopteron nebo Osteolepis. Na druhou stranu choana mohla také soužit k nějaké jiné funkci u ryb dýchajících žábrami. Kanálek vedoucí do očního důlku je porovnatelný se slzným kanálkem, který je nalézán u moderních tetrapodů. Sekrece ze žláz zvlhčuje oči a je odváděna přes nosní dutinu a chrání před poškozením očí, které jsou vystaveny volnému vzduchu. Podobnou funkci mohla dovolovat choana u lalokoploutvých ryb, které se nacházely blízko vodní hladiny. Dýchání přes choanu se mohlo vyvinout později. Choana se poprvé objevuje u druhu Kenichthys campbelli ze spodního devonu 3,4. U tohoto druhu se jedná o jakési přechodné stádium k pravé choaně (viz Obr. ).
Co na to genetika
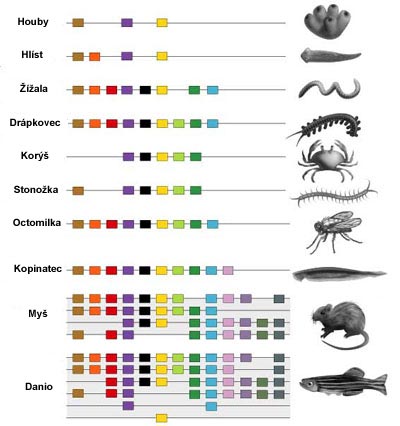 |
| Obr. 20 Hox geny u různých skupin živočichů |
Kromě fosilního materiálu nám o evoluci tetrapodů může mnohé říct i gnešní genetika. Díky genetice můžeme porozumět změnám, které ryby a jejich ploutve prodělaly, aby se z nich stala plnohodnotná končetina s prsty. Kde se vůbec končetiny a hlavně párové ploutve vzaly?
Rozdíl mezi prvními obratlovci a prvními bezobratlými nebyl ani tak velký. Můžeme si je představit jako jakési torpéda - válcovitý plán těla. Rozlišení jednotlivých úseků v předozadní ose řídí skupina tzv. homeotických (Hox) genů. Zatímco bezobratlí mají jen jednu sadu, tak obratlovci mají hned čtyři (a víc) sady těchto Hox genů. Toto se vysvětluje dvojitým zdvojením celé sady chromosomů.
Jak se zdá, tak velkou roli v evoluci ploutví a končetin hrály T-box geny (zkracovány také Tbx). Tyto geny byly objeveny terpve nedávno. Kódují proteiny, které se váží do regulačních oblastí jiných genů a tím vyvolávají jejich transkripci. Tyto geny se objevují už u nejprimitivnějších mnohobuněčných živočichů. Byly dokonce objeveny i u primitivního vločkovce rodu Trichoplax. Pro vznik končetin hrají zásadní roli T-box geny z rodiny T-box 2. Konkrétně se jedná o gen T-box 4 a T-box 5. T-box 5 jednoznačně určuje identitu přední končetiny - ať už je to prsní ploutev, přední končetina, křídlo nebo ruka. T-box 4 zase určuje to, že se bude jednat o břišní ploutev, zadní končetinu nebo nohu. Nejjednodušší strunatci, jako jsou kopinatci, ještě žádné končetiny ani ploutve nemají a stejně tak ani nemají žádný z těchto genů. U kopinatců však existuje jeden gen, který připomíná oba geny T-box 4 a T-box 5. Proto je také označován jako T-box 4/5. Duplikací genomu zřejmě došlo k duplikaci tohoto genu a dále k samostatné evoluci T-box 4 a T-box 5 genu. 6
 |
| Obr. 21 Kopinatec Branchiostoma |
Role T-box 4 a T-box 5 genů během "molekulární" fáze iniciace vzniku končetinového pupenu v povrchové vrstvě mezodermu ukazuje na to, že prvotní a jedinou rolí této dvojice genů nebylo specifikovat přední nebo zadní končetinu, ale zahajovat proces formování nějaké struktury vystupující bočně z povrchu torpédovitého těla. To byla asi původní úloha ještě nerozlišeného kopinatčího T-box 4/5 genu. Indukce vzniku ploutevního lemu dobře odpovídá takové představě. Docházelo k ní zřejmě v celém pásu podél obou boků. 6
 |
| Obr. 22 Placoderm Pterichthyodes |
Donedávna však předpokládanou roli genu T-box 4/5 při formování ploutevního lemu nepřímo dokládal právě jen strunatec (ne však obratlovec) kopinatec. A tak výzkumníky z takzvané disciplíny "Evo-Devo", která se snaží právě přes porozumění detailům procesů vývoje zárodku vysvětlit průběh evoluce živých organismů, rozdělovala otázka, nebo spíše pochybnost, jestli ploutevní lem byl skutečně mezistupněm vzniku párových končetin. Paleontologické nálezy fosilií prvních obratlovců totiž poskytovaly trošku odlišný obraz: zástupci vyhynulého kmene obratlovců - pancířnatých (Placodermi) mají totiž vyvinutý jen přední pár ploutví, který nasedá na lebku. To by tedy mohlo ukazovat na jiný průběh událostí, v rámci kterého nejprve vznikl v hlavové části jeden pár ploutví, který byl během dalšího vývoje jako již hotová struktura duplikován do zadní části těla, kde se objevil jakožto břišní ploutve. 6
Na druhou stranu jiný vyhynulý kmen obratlovců - trnoploutví (Acanthodii) z období siluru až permu, s řadou párových ploutví (ovšem poněkud menších) mezi párem prsních a párem břišních ploutví, by mohl být jakousi přechodnou formou mezi ploutevním lemem a dvěma páry ploutví. To by zapadalo do scénáře, podle kterého nediferencovaný gen T-box 4/5 kódoval vznik ploutevního lemu a po rozdělení na T-box 4 a T-box 5 mohl začít proces postupného rozrůzňování jejich senzitivity vůči předo-zadní poziční informaci, vytvářené homeotickými geny ve svrchní vrstvě mezodermu. 6
 |
| Obr. 23 Myllokunmingia - rekonstrukce |
Nesoulad mezi "Evo-Devo" a paleontologií nakonec smazala sama paleontologie. Tím novým podnětem je objev dvou fosilií z takzvané Cheng-jiangské fauny, pojmenované podle naleziště v jihočínské provincii Ju-nan. Myllokunmingia fengjiaoa a Haikouichthys ercaicunensis jsou obratlovci rybovitého (torpédovitého) tvaru těla, zařazovaní mezi bezčelistnaté obratlovce (Agnatha) (skupina, kam patří mihule a mořské sliznatky) nebo alternativně, ale s určitými pochybnostmi, k chimérám – což je skupina druhů velice bizarních rybovitých obratlovců ze třídy paryby. Cheng-jiangská fauna je datována do období kambria, je tedy daleko starší, než byly nálezy prvních doposud popsaných obratlovců. Podstatné je, že obě fosílie mají ploutevní lem. 6
Kromě těchto dvou T-box genů samozřejmě hrají při embryogenezi končetin roli také další regulační geny. Nedávný výzkum provedený se žraloky ukázal, že za vývojem ploutví u tak primitivních obratlovců jako jsou paryby stojí ty samé regulační geny, které hrají důležitou roli i při vývoji končetin u tetrapodů. Jedná se o regulační geny HoxD, Fgf8 a Tbx18.
Pět prstů
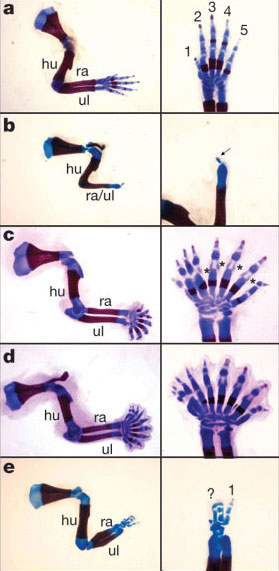 |
| Obr. 24 Ukázka exprese genů Shh a Gli3 8 |
Dříve se myslelo, že prsty vznikly z radiálií ploutví (např. viz Sauripteris Obr.) a jsou k nim homologické. Tuto představu však narušily první objevy primitivních obojživelníků, kteří měli osm, sedm nebo šest prstů. Do té doby se totiž myslelo, že pětiprstý plán končetiny je původní. Další výzkumy a objevy však tuto teorii zcela vyvrátily. Dnes se domníváme, že vývoj šel přes zánik radiálií až po vznik nových postaxiálních elementů.
Nedávno se ukázalo, že pětiprstou končetinu mají na svědomí regulační geny. Jedná se o již zmíněný gen Sonic hedgehog (dále jen Shh) a gen Gli3.
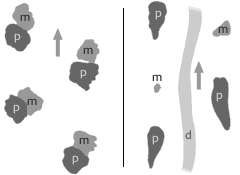
Na obrázku a) je vidět přední končetina myši s oběma aktivními geny Shh a Gli3. Tyto geny jsou aktivní při normálním embryonálním vývoji a vzniká tedy normální pětiprstá končetina s normálním stylopodem, zeugopodem a autopodem.
Na dalším obrázku b) je vidět výsledek embryonálního vývoje při absenci genu Shh. Vzniklá končetina má normální stylopodum, redukované nebo spojené zeugopodum a neidentifikovatelný element místo autopoda.
Další obrázek c) ukazuje končetinu, kde byl při vývinu aktivní pouze Shh a potlačen byl gen Gli3. Výsledkem je prakticky normální končetina, ale místo pěti prstů jich je víc - mezi šesti až jedenácti.
Další obrázek d) ukazuje situaci, kdy jsou neaktivní oba geny Shh a Gli3. Výsledek je podobný situaci na obrázku c) s tím rozdílem, že prsty postrádají identitu. Jinými slovy jsou všechny prsty stejné.
Na posledním obrázku e) je situace s pouze jednou kopií genu Gli3 a neaktivním genem Shh. Výsledkem je téměř kompletní končetina ovšem jen s několika prsty, které lze identifikovat jako prst 1.
Právě z obrázku c) a d) můžeme vidět to, jak asi působily geny na končetinu u druhů jako byla Acanthostega, Ichthyostega nebo Tulerpeton. Právě výzkum genů Shh a Gli3 spolu s fosilními doklady nám dávají ucelený obraz o vývoji pětiprsté končetiny. Díky tomuto výzkumu jsme mohli dostat odpověď na otázku polydaktilních končetin u primitivních obojživelníků.
Závěr
Z výše uvedeného je vidět, že doklady o vývoji končetiny z ploutve jsou nejen z oblasti paleontologie, ale také z oblasti genetiky. Zatímco genetika nám dává porozumět dějům, které probíhají na molekulární úrovni, tak paleontologie nám ukazuje skutečnou cestu, kterou se vývoj ubíral.
[1] Murphy, Dennis C. Devonian Times
[2] Boisvert, Catherine A. The pelvic fin and girdle of Panderichthys and the origin of tetrapod locomotion. Nature 438. 2005
[3] Evolutionary Ecology: The Devonian Opportunity
[4] Zhu, Mai, Ahlberg, Per E. The origin of the internal nostril of tetrapods. Nature 432. 2004
[5] Michael I. Coates, Martin J. Cohn. Fins, limbs, and tails: outgrowths and axial patterning in vertebrate evolution. 1998
[6] Hanák, P. Jak jsme přišli k rukám a nohám
[7] Chin Chiang, Ying Litingtung. VUMC team pinpoints genes that dictate five fingers. 2002
[8] Litingtung, Y., Dahn, R. D., Li, Y., Fallon, J. F., Chiang, Ch. Shh and Gli3 are dispensable for limb skeleton formation but regulate digit number and identity. Nature 418, 979-983. 2002
[9] UF scientists discover evolutionary origin of fins, limbs